1. La realidad biológica de Chrysodeixis includens: un comportamiento larvario que rompe el modelo tradicional
Para cualquier asesor técnico o agricultor, la escena en campo se ha vuelto frustrantemente familiar: tras realizar una aplicación fitosanitaria rigurosa contra lepidópteros defoliadores, el muestreo a las 48 horas revela que las poblaciones de la «falsa medidora» o «oruga agrimensora» (Chrysodeixis includens) permanecen prácticamente inalteradas. Lo que inicialmente se detecta como pequeños daños dispersos en el tercio inferior del lote, puede transformarse en cuestión de días en una defoliación severa que compromete directamente el rendimiento fotosintético de plantas de gran valor económico como la soja, el algodón, el girasol, el tomate y el tabaco
Técnicos y agricultores esperan observar las señales habituales para realizar los tratamientos fitosanitarios. Pero el monitoreo diurno tradicional muestra un número de orugas engañosamente bajo, haciendo que se subestime la población real. No se ven muchas larvas porque a partir de L3 las larvas regulan su ritmo circadiano para proteger su metabolismo. Durante el día se ocultan en el envés de las hojas y zonas sombreadas para evitar la radiación UV y la desecación, activándose solo de noche. Además el cultivo parece estar sano y estable durante días, pero de un momento a otro sufre pérdidas foliares drásticas y destructivas. Lo que sucede es que en un plazo críticamente corto las larvas pasan del estadio L2 a L5, cuando se empiezan a activar enzimas hidrolíticas que la llevan a consumir hasta diez veces más tejido que una L2.
¿Sabes EXACTAMENTE cuándo eclosiona la L1 en tu lote hoy?
Deja de adivinar y desbloquea el Seguimiento Fenológico en Tiempo Real de Ch. includens. Recibe alertas automáticas basadas en Grados Día de tu zona y encuentra la ventana perfecta para un control de alta eficacia.
Comienza tu Prueba Gratuita de 7 Días
La presencia de la plaga es constante y simultánea, de larvas de todos los tamaños (grandes y pequeñas al mismo tiempo), incluso pocos días después de haber realizado una aplicación. Además parece que los tratamientos «funcionan a medias»: Se tiene la percepción de que los insecticidas aplicados son ineficaces o de mala calidad debido a que la población no disminuye como se esperaba. En realidad, a partir de L3 se activa un escudo bioquímico por la sobreexpresión interna de citocromos P450, glutatión-S-transferasas (GST) y esterasa, familias enzimáticas que destruyen y metabolizan la molécula del insecticida antes de que toque sus órganos diana.
Por estas razones, hoy en día Chrysodeixis includens es uno de los defoliadores más destructivos del continente americano. A pesar de su impacto, el control en campo sigue basándose, en la mayoría de los casos, en un principio operativo tradicional y reactivo: aplicar insecticidas cuando se observa daño foliar visible o se detectan larvas en el muestreo.
Este enfoque de «veo daño -> trato» ha funcionado razonablemente bien durante décadas para un gran número de especies (como Helicoverpa, Spodoptera frugiperda o Rachiplusia nu) que poseen ciclos relativamente definidos, un comportamiento alimentario estable y una alta exposición en el follaje tratado. De hecho, las herramientas químicas del mercado están formuladas para esa realidad biológica lineal: productos de contacto para larvas expuestas, ingestivos para larvas activas y reguladores de crecimiento (IGR) para estadios tempranos. Sin embargo, esta lógica solo es válida cuando el desarrollo de la plaga es predecible y homogéneo.
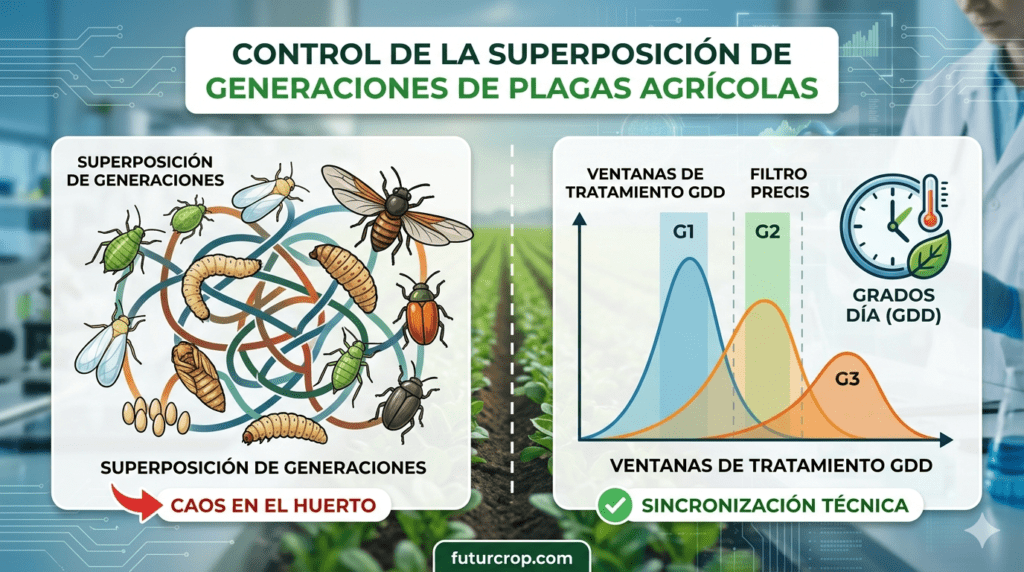
Con la falsa medidora, nos encontramos ante una biología que desborda por completo el modelo químico tradicional. La plaga combina una altísima capacidad defoliadora con una tolerancia creciente a las moléculas químicas. Sus cambios metabólicos bruscos entre estadios, su actividad estrictamente nocturna, su temprana capacidad detoxificadora y su hábito de refugio profundo en el dosel foliar provocan un fenómeno crítico: cuando el daño se hace evidente a simple vista, la ventana de máxima eficacia fitosanitaria ya ha pasado de largo.
Por qué los insecticidas fallan con Chrysodeixis includens
La industria formuló insecticidas para un modelo de oruga que C. includens no cumple.
1. Un modelo de insecticida que C. includens no respeta
La industria formuló insecticidas asumiendo orugas: expuestas en el follaje, con ciclos relativamente sincronizados, alimentación continua y alta susceptibilidad en estadios tempranos. Este modelo funciona para Helicoverpa, Spodoptera o Rachiplusia, pero no para Chrysodeixis includens.
En C. includens la biología rompe esos supuestos: oviposición continua (L1–L6 simultáneos), cambio brusco de comportamiento entre L2 y L3, refugio diurno en el envés, actividad nocturna y una capacidad detoxificadora muy superior. El resultado es que los tratamientos “correctos” en papel no se traducen en control eficaz en campo.
Los productos no son necesariamente malos; simplemente fueron diseñados para una oruga que está expuesta, sincronizada y con menor capacidad metabólica. C. includens hace exactamente lo contrario: se refugia, se desincroniza y activa mecanismos de defensa fisiológica a partir de L3.
2. Especies con comportamiento “estándar” vs “no estándar”
| Especie | Tipo de comportamiento | Características clave | Consecuencias para el control |
|---|---|---|---|
| Helicoverpa armigera | Estándar | Alimentación continua, alta exposición, ciclos definidos. | El manejo reactivo suele funcionar; buena respuesta a insecticidas. |
| Spodoptera frugiperda | Estándar con variaciones | Larvas móviles pero visibles; daño progresivo. | Control aceptable si se interviene temprano. |
| Rachiplusia nu | Estándar | Comportamiento alimentario estable; refugio limitado. | Buena eficacia de IGR y productos de contacto. |
| Chrysodeixis includens | No estándar | Cambio brusco entre L2 y L3; actividad nocturna; refugio; detoxificación elevada. | El manejo reactivo falla; requiere predicción fenológica y sincronización con L1–L2. |
| Tuta absoluta | No estándar | Penetración en tejido; refugio interno. | Control difícil sin predicción y estrategias específicas. |
| Tecia solanivora | No estándar | Larvas internas desde el inicio. | Tratamientos foliares prácticamente ineficaces. |
3. Eficacia de insecticidas en C. includens frente a otras orugas
Piretroides
En C. includens la mortalidad con λ‑cyhalothrin o bifentrina es baja si no se usan sinergistas. La necesidad de PBO o DEM confirma una resistencia metabólica marcada.
Frente a Spodoptera o Helicoverpa, la susceptibilidad es claramente menor, lo que obliga a aumentar dosis, mezclar productos o repetir aplicaciones.
Resistencia metabólica
Estudios transcriptómicos muestran hasta 28,9× más resistencia a λ‑cyhalothrin entre cepas resistentes y susceptibles, con sobreexpresión de múltiples P450, GST, esterasas y UGT.
El nivel de sobreexpresión es superior al observado en otras orugas bajo condiciones similares, lo que explica por qué C. includens “aguanta” mejor los tratamientos.
Diamidas
En Brasil se ha documentado una tolerancia elevada a ciantraniliprole y resistencias de hasta 70× a flubendiamida en poblaciones de campo.
Mientras que otras especies siguen siendo bien controladas por diamidas, en C. includens la eficacia se erosiona rápidamente si no se acompaña de manejo integrado y rotación real de modos de acción.
Síntesis biológica
Chrysodeixis includens es más difícil de controlar que otras orugas porque combina: detoxificación metabólica extraordinaria, tolerancia elevada a diamidas, resistencia emergente, comportamiento nocturno y refugio, y superposición generacional.
No es una plaga “mal controlada” por falta de productos, sino una plaga cuya biología desborda el modelo químico reactivo. El control eficaz exige anticipación, predicción fenológica y sincronización con los estadios más vulnerables (L1–L2).
Este escenario evidencia un cambio de paradigma crítico en la protección vegetal moderna. Durante décadas, el control de plagas ha descansado sobre un modelo puramente químico, caracterizado por tratamientos pautados rígidamente por calendario o activados tras la observación visual de umbrales económicos de daño cuando la plaga ya es madura. Sin embargo, Chrysodeixis includens ha demostrado de manera contundente que este enfoque está obsoleto. Su particular biología y comportamiento no solo eluden la acción de los ingredientes activos más comunes, sino que convierten el uso mal planificado de agroquímicos en un factor que agrava el problema en lugar de solucionarlo.
2. El Impacto Económico y la Evolución Histórica de la Plaga
Originalmente considerada una plaga secundaria en muchas regiones agrícolas de América (desde el sur de Estados Unidos hasta las pampas argentinas), Chrysodeixis includens ha escalado posiciones hasta convertirse en uno de los artrópodos fitófagos más costosos y destructivos del continente. En cultivos clave como la soja, su capacidad para alimentarse intensamente del parénquima foliar respetando únicamente las nervaduras (dejando la hoja con un aspecto característico de «encaje») genera pérdidas directas millonarias debido a la reducción del llenado de granos.
La evolución histórica de este problema está ligada intrínsecamente a la intensificación agrícola. La adopción masiva de monocultivos, la introducción de variedades transgénicas que controlan de forma selectiva solo a ciertas especies de lepidópteros (dejando nichos ecológicos vacíos) y, sobre todo, el uso indiscriminado de insecticidas de amplio espectro para controlar otras plagas concurrentes, actuaron como detonantes. Al eliminar sistemáticamente a la fauna benéfica —predadores como chinches pentatómidos y parasitoides de las familias Trichogrammatidae y Braconidae—, se rompió el control biológico natural de la falsa medidora. Paralelamente, la enorme presión de selección química aceleró el desarrollo de poblaciones con una resistencia genética extraordinaria, transformando un insecto secundario en una amenaza primaria de difícil gestión.
Nota de Campo: El coste de control de Chrysodeixis includens en regiones de alta presión fitosanitaria se ha triplicado en la última década, obligando a los productores a realizar hasta 4 o 5 aplicaciones específicas por ciclo de cultivo con moléculas de última generación, elevando drásticamente los costes de producción y reduciendo los márgenes de rentabilidad.
Evolución histórica del control
En los años 90, C. includens era una plaga secundaria. Los piretroides funcionaban bien porque las poblaciones eran menos resistentes y los sistemas de producción menos intensivos.
A partir de los 2000, con la expansión de la soja en Brasil y Argentina, la plaga encontró un ambiente ideal: temperaturas altas, disponibilidad continua de hospedantes y un uso intensivo de insecticidas que aceleró la selección de resistencia.
En la década de 2010, los piretroides dejaron de funcionar de manera consistente, las diamidas comenzaron a mostrar tolerancia creciente y los productores empezaron a aplicar mezclas cada vez más complejas. En México, por ejemplo, los técnicos reportan que dos aplicaciones consecutivas de diamidas ya no garantizan el control, especialmente en zonas cálidas donde la plaga completa su ciclo en apenas 18–22 días.
Hoy, el control químico es más caro, menos eficaz y más frecuente que nunca. Y sin embargo, la plaga sigue expandiéndose.
Evolución Anual del Coste y Pérdidas Económicas
Tendencia histórica combinada de Chrysodeixis includens (falsa medidora). Visualización de costes de manejo (líneas gruesas) y pérdidas anuales totales estimadas por país (líneas finas discontinuas).
El gráfico de Manejo Reactivo (Coste Alto, Eficacia Baja) muestra que un mayor gasto acumulado enUSD/ha ocurre cuando el control se realiza tarde. El incremento en el gasto se debe principalmente a un «Rescate Químico Múltiple».
Este gasto elevado no disminuye las pérdidas debido a la biología de la plaga. Al momento del «Muestreo Visual Típico» (etapas L3-L4), las orugas maduras tienen una cutícula más gruesa y una detoxificación 20 veces mayor. Además, estas orugas se refugian en la base del cultivo (L5-L6), lo que resulta en eficacia baja del tratamiento químico. Como consecuencia, las orugas siguen alimentándose, provocando una «Pérdida de Rendimiento» que se traduce en altas pérdidas económicas (USD/Año).
La situación donde el que más gasta es el que más sigue perdiendo se ilustra claramente en el gráfico de Pérdidas Anuales Brasil (MM USD). Brasil gasta la mayor cantidad de USD/ha en manejo reactivo, lo que se refleja en una línea roja que alcanza un pico de costes de control de más de 90 USD/ha al final del ciclo. Sin embargo, la línea verde de pérdidas anuales también alcanza un pico máximo de más de 300 MM USD/Año para Brasil. Esto demuestra que gastar grandes sumas en control reactivo y tardío no logra reducir las pérdidas.
Impacto económico real en América
Pérdidas económicas por infestación de Chrysodeixis includens
En soja, los estudios del USDA y del Embrapa (Brasil) estiman pérdidas de hasta 30–40 % del rendimiento en infestaciones severas sin control.
En condiciones de alta densidad larvaria (más de 30 larvas/m²), se han registrado reducciones de 600–900 kg/ha en rendimiento de grano.
En el sur de Brasil y norte de Argentina, los daños acumulados por C. includens representan entre 15 % y 25 % de las pérdidas totales por defoliadores, superando a Rachiplusia nu en años cálidos.
En el sur de EE. UU., el costo estimado de pérdidas directas y control supera los US $ 200 millones anuales en soja, según reportes de la Soybean Looper Working Group (2023).
Gasto medio en insecticidas por hectárea
En Brasil, los productores destinan entre 80 y 90 USD/ha solo a insecticidas contra esta plaga. En Argentina, el gasto oscila entre 60 y 75 USD/ha, mientras que en Paraguay ronda los 50–60 USD/ha.
En México, especialmente en Sinaloa, Tamaulipas y Veracruz, el coste medio se sitúa entre 45 y 60 USD/ha, dependiendo del cultivo y del número de aplicaciones.
En el sur de Estados Unidos, donde la presión es menor pero constante, el gasto ronda los 45–55 USD/ha.
Si se multiplica este gasto por la superficie afectada —más de 50 millones de hectáreas en América—, el impacto económico anual supera fácilmente los 2.000 millones de dólares. Y esto sin contar las pérdidas de rendimiento, que pueden alcanzar el 20–40% en situaciones de alta presión.
Correlación entre gasto en insecticidas y pérdidas por Chrysodeixis includens
El siguiente gráfico muestra cómo las regiones con mayor gasto medio en insecticidas por hectárea son también las que sufren mayores pérdidas potenciales si no se controla la plaga. La correlación es clara: mayor presión → mayor gasto → mayores pérdidas.
La línea de tendencia muestra que las regiones con mayor gasto (Brasil, Argentina) son también las que presentan mayores pérdidas potenciales. Esto confirma que el modelo reactivo basado en aplicaciones tardías es ineficiente frente a una plaga con superposición generacional, refugio diurno y resistencia metabólica.
Existe una correlación positiva entre el gasto medio en insecticidas y las pérdidas potenciales sin control. Las regiones con mayor presión de C. includens (Brasil, Argentina) muestran tanto mayor gasto como mayores pérdidas, lo que refleja la dificultad de lograr un control eficaz con estrategias tradicionales.
Por qué Chrysodeixis includens genera el mayor gasto en control
- Superposición generacional constante La oviposición continua produce poblaciones con larvas de todos los estadios simultáneamente. Esto obliga a realizar múltiples aplicaciones para cubrir cohortes sucesivas.
- Resistencia metabólica y fisiológica La sobreexpresión de enzimas detoxificadoras (P450, GST, esterasas, UGT) reduce la eficacia de los insecticidas, especialmente piretroides y diamidas. Resultado: más dosis, más mezclas, más gasto.
- Comportamiento nocturno y refugio diurno Las larvas se alimentan principalmente de noche y se ocultan durante el día en el envés de las hojas. Esto disminuye el contacto con el insecticida y exige productos de mayor persistencia o penetración.
- Defoliación rápida y detección tardía El salto entre L2 y L3 provoca un aumento exponencial del consumo foliar. Cuando el técnico detecta el daño, la ventana óptima de control (L1–L2) ya ha pasado. Resultado: tratamientos tardíos y repetidos.
- Dependencia del control químico En la mayoría de regiones, el manejo sigue siendo reactivo y basado en insecticidas. La falta de integración de modelos fenológicos y monitoreo predictivo mantiene el gasto elevado.

3. La Explicación Científica: Por Qué la Plaga Rompe el Modelo Químico
El fracaso recurrente de las intervenciones químicas tradicionales no se debe a una mala calidad de los productos comerciales, sino a mecanismos biológicos, enzimáticos y etológicos sumamente eficientes que posee el insecto.
3.1. El arsenal invisible de la detoxificación metabólica
A nivel fisiológico, las larvas de Chrysodeixis includens poseen una de las maquinarias enzimáticas más complejas del mundo de los insectos, diseñada para neutralizar compuestos tóxicos exógenos (tanto defensas naturales de las plantas como insecticidas sintéticos). Estudios genómicos y bioquímicos han mapeado este sofisticado «ejército» de detoxificación, destacando los siguientes componentes:
- 15 Citocromos P450 (Monooxigenasas): Superfamilia de enzimas que alteran la estructura molecular del insecticida mediante procesos de oxidación, reduciendo su toxicidad drásticamente antes de que alcance los sitios diana en el sistema nervioso o muscular del insecto.
- 7 Glutatión-S-Transferasas (GST): Enzimas que catalizan la conjugación de moléculas fitosanitarias con compuestos endógenos, volviéndolas hidrosolubles y facilitando su rápida excreción fuera del organismo de la larva.
- 7 Esterasas: Especializadas en la hidrólisis de enlaces éster presentes en una amplísima gama de insecticidas tradicionales (como organofosforados, carbamatos y piretroides), inactivando el principio activo con extrema velocidad.
- 8 UDP-Glucuronosiltransferasas (UGT): Complejos enzimáticos que unen azúcares a las moléculas hidrofóbicas exógenas, completando el proceso de desactivación metabólica e impidiendo la bioacumulación del tóxico.
3.2. Comportamiento etológico defensivo
Además de su blindaje químico interno, la falsa medidora despliega un comportamiento que dificulta físicamente el contacto con las pulverizaciones. Las hembras adultas realizan la oviposición de forma aislada, preferentemente en el envés de las hojas situadas en el tercio inferior y medio de la planta. Al eclosionar, las larvas se alimentan estrictamente en esa zona interna y sombreada del dosel foliar. Esto genera un «efecto paraguas»: las hojas superiores actúan como una barrera física que retiene la mayor parte de las gotas del insecticida, permitiendo que solo una fracción mínima del producto llegue al estrato inferior donde la plaga está activa.
3.3. Superposición generacional
Debido a su ciclo biológico fuertemente influenciado por las condiciones microclimáticas, es habitual encontrar en un mismo momento del cultivo huevos, larvas de múltiples tamaños (desde L1 a L6), pupas y adultos conviviendo simultáneamente. Un tratamiento químico convencional con una única molécula solo afectará a una fracción de la población, dejando intactos los demás estadios y provocando continuas «olas» de reinfestación pocos días después de la aplicación.
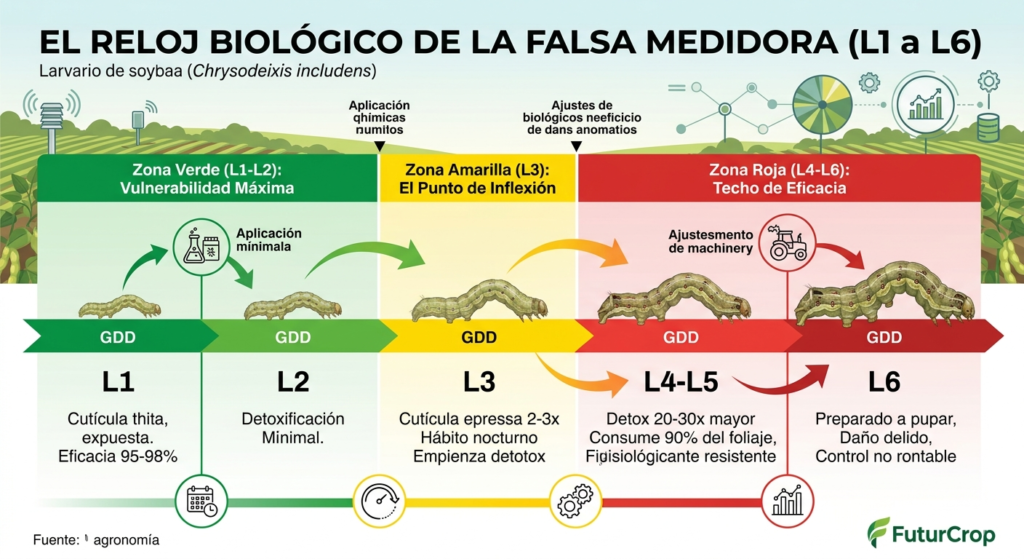
El desarrollo larvario no estándar
Los estadios L1 y L2 duran entre 4 y 6 días. Durante este periodo, la larva es extremadamente vulnerable: su cutícula es fina, su capacidad detoxificadora es mínima y permanece expuesta en la superficie de la hoja. Sin embargo, el daño que causa es tan leve que pasa desapercibido para el técnico.
El salto entre L2 y L3 es el punto crítico. En apenas 24–36 horas, la larva experimenta un aumento brusco de tamaño, su aparato bucal se fortalece, su movilidad aumenta y su comportamiento cambia por completo. Este cambio está regulado por un pico de ecdisona, la hormona que desencadena la muda. La ecdisona activa la expresión de genes asociados al engrosamiento de la cutícula, la maduración del sistema digestivo y la activación de enzimas detoxificadoras.
A partir de L3, la larva se vuelve predominantemente nocturna. Durante el día se refugia en el envés de las hojas o en zonas sombreadas del dosel, donde la humedad es mayor y la radiación UV es menor. Este comportamiento reduce drásticamente la exposición al insecticida.
Los estadios L4, L5 y L6 duran entre 2 y 4 días cada uno, dependiendo de la temperatura. En esta fase, la larva presenta una capacidad detoxificadora plenamente desarrollada, una cutícula gruesa y una tasa de consumo explosiva. Una larva L5 puede consumir 10 veces más tejido que una L2.

4. Radiografía por Estadios Larvarios: Vulnerabilidad vs. Resistencia
El ciclo larvario de Chrysodeixis includens suele comprender entre 6 estadios (L1 a L6), cada uno con características morfológicas, fisiológicas y conductuales radicalmente diferentes. Entender esta progresión es la clave absoluta para un control eficaz.
| Estadio Larvario | Características Biológicas y Conductuales | Nivel de Daño | Vulnerabilidad Química |
| L1 – L2 (Instars Iniciales) | Larvas de tamaño reducido (2-5 mm). Cutícula extremadamente fina y permeable. Su aparato enzimático defensivo está inmaduro y apenas expresan los genes de detoxificación metabólica. Se localizan expuestas de forma más externa. | Mínimo (Pequeños raspados foliares casi imperceptibles). | 🟢 MÁXIMA (Ventana crítica de control eficaz). |
| L3 (Transición) | La larva incrementa su tasa de consumo. Comienza a migrar hacia el interior profundo del dosel foliar. El sistema endocrino empieza a activar la producción de citocromos y esterasas de defensa. | Moderado (Inicio de perforaciones pequeñas). | 🟡 MEDIA (Requiere dosis completas y buena cobertura). |
| L4 – L6 (Instars Maduros) | Larvas grandes (hasta 35 mm) con movimientos típicos de bucle o «medidores». Cutícula gruesa y quitinizada. Los 15 citocromos P450, las GST y esterasas están sobreexpresados y funcionando al 100%. Consumen más del 90% del total de biomasa foliar de todo su ciclo. | 🔴 CRÍTICO (Gran capacidad de defoliación en 48 horas). | 🔴 MÍNIMA / NULA (Prácticamente indestructibles por vía química). |
Como demuestra la tabla, se produce una paradoja agronómica perversa: el agricultor suele alarmarse y decidir la aplicación química cuando observa defoliaciones severas en el campo, es decir, cuando la población está dominada por estadios L4-L6. En ese momento, intentar el control químico es agronómicamente inútil, económicamente ruinoso y un acelerador directo de la selección de resistencias.
5. El Fracaso del Modelo Químico Convencional
Los técnicos aplican insecticidas cuando ven daño. Pero cuando el daño es visible, la población ya está dominada por L4–L6, los estadios más resistentes y menos expuestos.
Eficacia de insecticidas en Chrysodeixis includens
Comparación con otras orugas agrícolas: un problema fisiológico, metabólico y de comportamiento.
Los estudios científicos coinciden en que Chrysodeixis includens es más difícil de controlar que otras orugas agrícolas como Spodoptera o Helicoverpa. La causa no es solo el producto, sino la combinación de baja susceptibilidad, resistencia metabólica muy elevada y tolerancia creciente a diamidas.
1. Baja susceptibilidad a piretroides
Ensayos con λ‑cyhalothrin y bifentrina muestran una mortalidad baja en C. includens. La eficacia aumenta de forma clara solo cuando se añaden sinergistas como PBO o DEM.
Esto indica que la plaga no es “insensible” al ingrediente activo, sino que presenta una resistencia metabólica basada en mecanismos de detoxificación que neutralizan el insecticida antes de que actúe.
2. Resistencia metabólica muy superior
Un estudio transcriptómico comparó una cepa resistente con una susceptible y encontró hasta 28,9× más resistencia a λ‑cyhalothrin.
Se observó sobreexpresión de:
- 15 citocromos P450
- 7 GST
- 7 esterasas
- 8 UGT
Este perfil de sobreexpresión es mucho mayor que el descrito para Spodoptera o Helicoverpa en condiciones similares, lo que explica por qué C. includens “aguanta” mejor los tratamientos estándar.
3. Tolerancia elevada a diamidas
Estudios brasileños han documentado una tolerancia significativamente mayor a ciantraniliprole y poblaciones de campo más tolerantes que las cepas susceptibles de referencia.
En el caso de flubendiamida, se han registrado niveles de resistencia de hasta 70× en poblaciones resistentes.
Mientras otras orugas siguen respondiendo bien a las diamidas, en C. includens la eficacia se erosiona rápidamente si no se acompaña de un manejo integrado y una rotación real de modos de acción.
Síntesis: una plaga diseñada para sobrevivir al modelo químico estándar
Chrysodeixis includens no es solo “otra oruga difícil”. Combina baja susceptibilidad a piretroides, una resistencia metabólica extraordinaria y una tolerancia elevada a diamidas. Frente a especies como Spodoptera o Helicoverpa, su perfil fisiológico y metabólico la convierte en una plaga estructuralmente más resistente al modelo químico tradicional, lo que obliga a replantear el manejo hacia estrategias basadas en predicción, sincronización y reducción de presión de selección.
Por qué la industria no resuelve el problema aunque lo conoce desde hace décadas
La industria fitosanitaria lleva más de veinte años estudiando el caso de C. includens. Sus departamentos de I+D han documentado la superposición generacional, la detoxificación metabólica, la nocturnidad, el refugio diurno y la erosión progresiva de eficacia de los insecticidas. No es un problema desconocido: es un problema diagnosticado.
Sin embargo, la solución real —la predicción fenológica y la sincronización con L1–L2— no forma parte de su modelo de negocio. La industria vende moléculas, no decisiones. Un sistema que reduce aplicaciones no es comercialmente atractivo para empresas cuyo crecimiento depende del volumen de ventas. Por eso, aunque conocen la causa del problema, su respuesta se limita a nuevas formulaciones, nuevas mezclas y nuevas campañas comerciales, pero no a cambiar el modelo.
En los últimos años han aparecido campañas que prometen “control total”, “nueva generación de diamidas”, “penetración superior”, “doble modo de acción”, pero ninguna aborda el verdadero problema: la biología de la plaga no coincide con el modelo químico para el que fueron diseñados los insecticidas. Las líneas de investigación actuales —microencapsulados, formulaciones con coadyuvantes, mezclas sinérgicas— mejoran el martillo, pero no cambian el hecho de que el problema no es un clavo.
6. La Solución: Gestión Fitosanitaria Basada en la Biología Predictiva
Aplicar el conocimiento biológico de C. includens al manejo diario implica ajustar tanto el monitoreo como la estrategia de intervención. El muestreo debe desplazarse hacia las zonas sombreadas y el envés de las hojas, que es donde las larvas se refugian durante el día. La sincronización de tratamientos con la aparición de L1–L2 es esencial, porque es el único momento en que los insecticidas —de contacto, ingestivos o reguladores de crecimiento— alcanzan su máxima eficacia.
Aquí los modelos fenológicos vuelven a ser determinantes. Permiten anticipar explosiones poblacionales antes de que sean visibles, estimar con precisión cuándo ocurrirá la eclosión masiva y evitar caer en aplicaciones tardías, cuando la población ya está dominada por L4–L6 y el margen de acción es mínimo. Sin esta predicción, el técnico siempre llega tarde; con ella, llega exactamente cuando debe.
El manejo también debe incluir la combinación de modos de acción para reducir la presión de selección y retrasar la resistencia, así como el monitoreo después de lluvias ligeras, que suelen favorecer la eclosión y modificar la dinámica poblacional.
Integrar estos elementos transforma el manejo de C. includens en un proceso más eficiente, más económico y más alineado con la biología real de la plaga. Es el paso necesario para dejar atrás el modelo reactivo y avanzar hacia un manejo verdaderamente predictivo, donde cada tratamiento se aplica cuando tiene sentido y se evita cuando es un desperdicio.
6.1. La ventana de tratamiento: por qué funciona. qué ocurre en L1–L2
La ventana de tratamiento de C. includens es breve, pero extraordinariamente eficaz. Comprende los estadios L1 y L2, que duran entre 4 y 6 días en total. En este periodo, la larva es biológicamente vulnerable por razones muy concretas.
6.1.1. La cutícula fina: la puerta abierta
En L1–L2, la cutícula es extremadamente delgada. La cutícula es la barrera física que determina cuánta molécula insecticida penetra en el cuerpo de la larva. En L1–L2, esta barrera es casi inexistente, lo que permite que los insecticidas de contacto penetren con facilidad, que los ingestivos alcancen rápidamente el sistema digestivo y que los reguladores de crecimiento interfieran eficazmente en la muda.
En L3, la cutícula se engrosa entre 2 y 3 veces, reduciendo la penetración de insecticidas en más del 60%.
6.1.2. Detoxificación mínima: la larva aún no sabe defenderse
La detoxificación metabólica es el proceso mediante el cual la larva neutraliza insecticidas. En L1–L2, la actividad de citocromos P450, GST y esterasas es muy baja. Esto significa que el insecticida llega intacto al órgano diana, la larva no puede metabolizarlo a tiempo y la mortalidad es alta incluso con dosis bajas.
En L3, la expresión de P450 aumenta entre 8 y 12 veces, y en L4–L5 puede aumentar hasta 20–30 veces.
6.1.3. Exposición total: la larva está donde está el residuo
En L1–L2, la larva se alimenta en la superficie de la hoja, sin refugio. Esto garantiza que entra en contacto directo con el residuo, ingiere tejido tratado y no evita la zona de aplicación.
En L3, la larva se vuelve nocturna y pasa el día en el envés, donde la deposición de insecticida es mínima.
6.1.4. Evidencia científica de la eficacia en L1–L2
Ensayos en Brasil, Argentina y EE. UU. muestran que:
- los IGR tienen 90–95% de eficacia en L1–L2,
- los piretroides superan el 80% en L1–L2 (pero <40% en L4–L6),
- las diamidas logran 95–98% en L1–L2, pero solo 60–70% en L5.
La ventana funciona porque la biología está de nuestro lado. Fuera de ella, la biología trabaja en nuestra contra.
6.2 Modelos Fenológicos y Grados Día (GDD): Ciencia Aplicada
Los modelos fenológicos en la agricultura son herramientas que predicen el desarrollo de una plaga (como Chrysodeixis includens) a lo largo de su ciclo vital (huevo, larva, pupa, adulto), basándose en la temperatura ambiente de la zona. Como los insectos son organismos ectotérmicos, su velocidad de desarrollo depende directamente del calor acumulado, lo que se calcula mediante la fórmula de Grados-Día Acumulados ($GDD$). Estos modelos permiten conocer matemáticamente en qué estadio se encuentra la plaga en cada momento, incluso antes de que los síntomas de daño sean visibles a simple vista.
Para el control eficaz de Chrysodeixis includens (falsa medidora), la predicción fenológica ofrece ventajas estratégicas que el muestreo visual tradicional no puede igualar:
- Anticipación de la «Zona Verde» (L1-L2): El software detecta con días de antelación el pico exacto de eclosión de los huevos. Esto permite al agricultor actuar preventivamente sobre las larvas pequeñas (estadios L1 y L2), justo en su único momento de máxima vulnerabilidad fisiológica, cuando la cutícula es fina, la detoxificación metabólica es mínima y la eficacia fitosanitaria roza el 98%.
- Ahorro de Tratamientos de Rescate (L4-L6): La predicción permite evitar la aplicación de insecticidas de «rescate», costosos e inútiles, sobre larvas maduras (L4 a L6), las cuales ya han causado el 90% del daño foliar, presentan una detoxificación enzimática 30 veces mayor y se encuentran refugiadas profundamente en el dosel, siendo fisiológicamente resistentes a la mayoría de las moléculas químicas.
- Optimización Operativa de la Maquinaria: Al predecir el estadio exacto de la plaga, el técnico puede coordinar la logística de aplicación, calibrar la maquinaria con la configuración de gotas adecuada (fina para cobertura en L1-L2, media para penetración en L3) y asegurar que cada euro invertido en sanidad vegetal alcance su máximo techo de rentabilidad.
Sincronización de Tratamientos: ¿Por qué fallamos en campo?
Muestreo Visual vs. Predicción Fenológica en el Lote de Soja.
Conclusión del Tablero
El muestreo tradicional (reactivo) se basa en síntomas tardíos. La predicción fenológica (proactiva) se basa en datos biológicos exactos. Sustituir la incertidumbre por información permite reducir aplicaciones y maximizar el ROI sanitario.
6.3. Estrategia de Rotación Inteligente de Modos de Acción (IRAC)
Disponer de la predicción fenológica precisa permite diseñar bloques de tratamiento inteligentes que protegen las tecnologías químicas disponibles y evitan la resistencia:
| Momento Biológico (FuturCrop) | Grupo Químico (IRAC) | Ejemplo de Ingrediente Activo | Justificación Técnica y Fisiológica |
| Pico de Eclosión de Huevos / Inicio L1-L2 | Grupo 15 (IGR – Inhibidores de Quitina) o Biológicos | Lufenurón / Bacillus thuringiensis / Baculovirus (ClbiNPV) | Larva con cutícula fina y alta ingesta. Máxima susceptibilidad a toxinas biológicas y alteradores de la muda. Impacto ambiental mínimo. |
| Consolidación de Estadios L2-L3 (Si persiste presión) | Grupo 22 (Inhibidores de canales de sodio) o Grupo 5 (Espinosinas) | Indoxacarbe / Espinetoram | Moléculas de alta eficiencia por ingestión, ideales para cortar la transición hacia estadios donde se dispara la detoxificación enzimática. |
| Estadios L4-L6 (Detectados por fallos ajenos al modelo) | Grupo 28 (Diamidas Antranílicas) | Clorantraniliprol / Broflanilida (Grupo 30) | Reservados estrictamente como herramientas de rescate extremo debido a su persistencia y acción sistémica / translaminar en el dosel. |
7.3. Optimización técnica de la pulverización
La sincronización biológica debe acompañarse de una correcta ingeniería de aplicación para vencer el «efecto paraguas» del dosel vegetal. Cuando el modelo fenológico indique el momento oportuno, se debe configurar el equipo de pulverización asegurando una presión constante, el uso de pastillas de cono hueco o con asistencia de aire que generen un espectro de gotas medianas-finas (capaces de penetrar las capas superiores de las hojas), combinando la mezcla con coadyuvantes tensioactivos y aceites vegetales metilados que optimicen la cobertura y la penetración en el envés foliar del tercio inferior.
Matriz de Control Eficaz de Chrysodeixis includens
Selecciona el estadio larvario actual detectado o previsto por FuturCrop para ver la estrategia óptima.
Sustento Científico Fitosanitario
La cutícula es extremadamente fina, lo que permite una penetración casi total del insecticida. La detoxificación metabólica es mínima: los P450, GST y esterasas están en niveles basales. La larva se alimenta en la superficie de la hoja, totalmente expuesta al residuo. Los IGR son especialmente eficaces porque interfieren en la síntesis de quitina justo cuando la larva depende de ella para crecer.
Notas clave para interpretar la matriz de control
Para un asesor técnico o un productor agrícola, esta tabla no debe leerse como una simple lista de productos comerciales, sino como una hoja de ruta dictada por la fisiología del insecto. A continuación, analizamos los cinco pilares biológicos que determinan el éxito o el fracaso rotundo en el campo:
1. La ventana L1–L2 no es una recomendación comercial: es un hecho biológico
Tratar la plaga en sus dos primeros estadios larvarios es la única garantía de éxito porque es el único momento del ciclo donde todas las variables juegan a favor del agricultor: la cutícula de la larva es extremadamente fina y permeable, su sistema de detoxificación metabólica está inactivo (niveles basales), y el insecto se alimenta expuesto en la superficie foliar. En esta ventana fitosanitaria, los reguladores de crecimiento (IGR) interrumpen con éxito la síntesis de quitina, los piretroides tradicionales aún ofrecen respuesta y las diamidas alcanzan su máxima eficacia (entre el 95% y el 98%). Fuera de esta ventana, cualquier caída en el control se debe a razones fisiológicas del insecto, no a un «mal producto».
2. La pérdida de eficacia en L3 no es gradual: es un colapso abrupto
Existe la falsa creencia de que la resistencia de la falsa medidora aumenta de forma escalonada día a día. La realidad es que el paso de L2 a L3 representa un salto evolutivo crítico y repentino. En cuestión de horas, la larva experimenta un pico de ecdisona que activa la sobreexpresión de los genes detoxificadores (Citocromos P450, GST, Esterasas y UGT) y engrosa su cutícula hasta tres veces. Además, cambia radicalmente su comportamiento hacia hábitos nocturnos, refugiándose en el envés profundo durante el día. Esta es la explicación científica de por qué un tratamiento aplicado con la misma dosis «que funcionaba perfectamente hace dos días», de repente deja de surtir efecto.
3. En estadios L4–L6, la biología del insecto destruye la molécula química
Cuando la población en el campo está dominada por orugas maduras, intentar un control químico convencional es agronómicamente inútil. No se trata de un problema de marcas comerciales, de sobredosificación o de formulaciones milagrosas; es fisiología pura. En esta fase, la capacidad de detoxificación de la larva es entre 20 y 30 veces mayor que en sus primeros días, su cutícula es prácticamente impermeable y su alimentación nocturna reduce al mínimo la exposición continuada a los residuos del fitosanitario. Las moléculas se metabolizan y se excretan antes de alcanzar el sistema nervioso o muscular del insecto.
4. La estrategia IRAC exige sincronización, no solo rotación de envases
El Comité de Acción contra la Resistencia a Insecticidas (IRAC) promueve la rotación de modos de acción, pero alternar productos a ciegas es ineficaz si se hace a destiempo. La verdadera presión de selección y el desarrollo de superresistencias ocurren cuando se aplica tarde (sobre poblaciones maduras con alta capacidad detoxificadora) o cuando se repite el mismo modo de acción sobre generaciones consecutivas del insecto. Para que la rotación de grupos químicos funcione, debe estar estrictamente sincronizada con los picos de eclosión previstos matemáticamente.
5. Por qué el muestreo visual tradicional siempre «llega tarde»
La correlación de datos de esta matriz expone el gran punto débil del monitoreo convencional en campo: el ojo humano. Por lo general, un operario o técnico detecta visualmente los daños por defoliación o la presencia de la plaga cuando las larvas ya han alcanzado el tamaño de los estadios L3 o L4. El agricultor toma la decisión de compra y aplicación justo en el momento exacto en que la curva de vulnerabilidad real de la plaga ha colapsado.
📈 Conclusión operativa: Romper este círculo vicioso requiere sustituir la reacción visual por la acción predictiva. Solo mediante el uso de modelos fenológicos y software de previsión como FuturCrop, que calcula los Grados Día ($GDD$) y anticipa la aparición masiva de los estadios L1-L2, es posible golpear a la plaga en su único momento de debilidad biológica, optimizando la inversión y asegurando la sanidad del cultivo.
7. Estrategia IRAC Y APLICACIÓN
7.1. Estrategia de secuencia inteligente: Rotación IRAC basada en ventanas fenológicas
Diseñar una estrategia de manejo de resistencias (IRM) según las directrices del Comité de Acción contra la Resistencia a Insecticidas (IRAC) no consiste en alternar productos al azar según las ofertas del mercado. Para Chrysodeixis includens, la rotación química debe estructurarse sincronizando el Modo de Acción (MoA) de la molécula con la susceptibilidad metabólica del estadio larvario previsto por el modelo predictivo:
- Primera generación, ventana crítica L1–L2 (Máxima vulnerabilidad): La estrategia arranca utilizando un Regulador del Crecimiento de los Insectos (IGR), como un inhibidor de la síntesis de quitina o un análogo de la hormona juvenil (Grupos IRAC 15, 16 o 18). En esta fase inicial, los IGR muestran su máximo techo de eficacia biológica, ya que interfieren directamente en los procesos de muda y deposición de la nueva cutícula, justo cuando la micro-larva depende críticamente de estos procesos bioquímicos para sobrevivir.
- Primera generación, transición hacia estadio L3: Si el software fenológico indica que un porcentaje de la población ha comenzado la transición hacia L3, la estrategia debe virar hacia una Diamida Antranílica (Grupo IRAC 28). Las diamidas activan los receptores de rianodina, provocando una liberación masiva e irreversible de calcio en las células musculares que causa parálisis y muerte por inanición. Su eficacia se mantiene alta en L3 (pese al inicio de la detoxificación enzimática), siempre y cuando se aseguren la dosis completa y un volumen de caldo óptimo.
- Segunda generación, nueva ventana L1–L2 (Corte del ciclo): Ante el nacimiento de la siguiente generación biológica, se prohíbe repetir los grupos anteriores para evitar la resistencia cruzada. El modelo dicta rotar hacia una Espinosina (Grupo IRAC 5). Al actuar sobre los receptores nicotínicos de la acetilcolina (un sitio diana del sistema nervioso completamente distinto al de las diamidas o IGR), se desactiva la presión de selección sobre los mecanismos metabólicos del insecto y se resguarda la vida útil de la tecnología química.
- Segunda generación, presencia de L3 (Control de escape): Para rescatar los individuos que escaparon y alcanzaron el tercer estadio, se recurre a un Inhibidor de la Respiración Mitocondrial (Grupo IRAC 21) o a un producto de alta eficiencia por ingestión con un modo de acción alternativo, bloqueando por completo la consolidación de la súper-defensa enzimática de la plaga.
⚠️ Principio Fitosanitario Clave: Esta secuencia no es una receta comercial fija ni un calendario rígido. Ilustra el principio fundamental de la agronomía moderna: el éxito no depende de elegir el producto más costoso del almacén, sino de ubicar la molécula correcta en el estadio biológico exacto mediante alertas predictivas.
7.2. Ingeniería de aplicación: Cómo adaptar la tecnología de pulverización al estadio larvario
La biología y el comportamiento etológico de la falsa medidora no solo dictan qué principio activo cargar en el tanque, sino cómo debe configurarse mecánicamente el equipo de aplicación. El comportamiento del insecto cambia drásticamente a medida que crece, y la tecnología de pulverización debe evolucionar con él:
En estadios L1–L2: Prioridad a la cobertura foliar uniforme
En los primeros instars, la larva es diminuta, tiene una cutícula permeable y se encuentra alimentándose en las zonas más externas y expuestas de las hojas.
- Estrategia técnica: El objetivo es la máxima cobertura superficial, no la fuerza de penetración profunda.
- Configuración en campo: Se recomiendan volúmenes de caldo moderados (entre 80 y 120 L/ha en aplicaciones terrestres) utilizando boquillas que generen un espectro de gotas finas a muy finas. Esto multiplica los impactos por centímetro cuadrado, asegurando que la larva ingiera o toque el residuo de inmediato. En esta etapa, la precisión matemática del momento de aplicación es infinitamente más importante que la agresividad física del tratamiento.
En estadio L3: Rompiendo el «efecto paraguas» del dosel
Al llegar al tercer estadio, la oruga activa su instinto fototrópico negativo: comienza a refugiarse profundamente en el interior de la masa foliar y a alimentarse preferentemente en el envés de las hojas durante las horas de menor radiación.
- Estrategia técnica: Vencer la barrera física de las hojas superiores (efecto paraguas) y lograr que las gotas penetren vertical y horizontalmente.
- Configuración en campo: Es obligatorio elevar los volúmenes de agua (120 a 150 L/ha), cambiar a boquillas de gota media (para evitar la deriva y la evaporación antes de llegar al estrato inferior) y ajustar la presión del equipo o el ángulo de pulverización (por ejemplo, boquillas de doble abanico). Aquí, un producto excelente aplicado con una mala cobertura técnica se traducirá inevitablemente en un fracaso de control.
En estadios L4–L6: El techo de la eficacia fitosanitaria
Cuando la plaga alcanza sus últimos instars maduros, se encuentra protegida de forma casi absoluta por su blindaje cuticular, su hiperactividad enzimática y su total resguardo diurno en la base del cultivo. En este punto, incluso una aplicación técnicamente perfecta con tecnología de punta tiene un techo de eficacia biológica sumamente bajo.
Frente a una población dominada por estadios L4-L6, la decisión más inteligente y rentable para el asesor agrícola ya no es evaluar «cómo calibrar mejor el equipo», sino determinar si económicamente tiene sentido realizar la aplicación. En la inmensa mayoría de los casos, la respuesta es negativa: el coste económico de los tratamientos de rescate con dosis máximas supera por amplio margen el beneficio real en el rendimiento, confirmando que el daño ya está hecho y que la ventana de control se ha cerrado por completo.
Estrategia Operativa Avanzada
Navega entre la rotación científica de materias activas y los requerimientos de ingeniería de pulverización en campo.
8. La biología del insecto como problema y como solución
La era de combatir las plagas mediante la fuerza bruta de la química continua ha llegado a su fin. La mayoría de los fallos de control en Chrysodeixis includens no se deben a productos deficientes, errores de dosis ni a la incompetencia en el campo. Tampoco es un problema exclusivo de la resistencia, aunque esta exista y agrave la situación. El verdadero motivo del fracaso fitosanitario es que el modelo tradicional de manejo —basado en la intuición o en el síntoma visual de «veo daño, luego aplico»— obliga a intervenir fuera de la ventana óptima, cuando la población ya está dominada por larvas grandes.
Chrysodeixis includens no es una plaga compleja por falta de herramientas, sino porque su comportamiento rompe por completo las reglas del juego de la pulverización convencional bajo cinco pilares biológicos:
- Falta de sincronía generacional: La superposición de cohortes hace que coexistan todos los tamaños de larvas a la vez.
- Brevedad e invisibilidad: La ventana biológica vulnerable (L1–L2) es sumamente estrecha y no se detecta a simple vista.
- Blindaje enzimático prematuro: La detoxificación metabólica se activa drásticamente a partir de L3.
- Comportamiento etológico de refugio: El hábito nocturno reduce de forma crítica la exposición física al residuo del insecticida.
- Defoliación exponencial concentrada: El daño severo ocurre en un periodo de tiempo muy corto, justo cuando la población ya ha entrado en estadios fisiológicamente resistentes.
En este escenario, los insecticidas funcionan —y con una eficiencia excelente— cuando impactan en L1–L2. Sin embargo, se desperdician por completo cuando se aplican en L4–L6. La diferencia real del éxito no se encuentra escrita en la etiqueta del envase, sino en el momento exacto de la aplicación.
9. La predicción fenológica: El verdadero salto tecnológico
Comprender el comportamiento larvario transforma la toma de decisiones en un proceso de ciencia aplicada que sustituye a la incertidumbre del lote. La verdadera innovación de la agricultura moderna no proviene del desarrollo constante de nuevas moléculas químicas —un modelo de negocio industrial diseñado para sostener el volumen de aplicaciones—, sino de la adopción de nuevas metodologías predictivas para decidir cuándo usarlas.
Aquí es donde los modelos fenológicos se convierten en la pieza central y obligatoria del manejo integrado de plagas (MIP). Dado que la biología de los insectos está gobernada rigurosamente por la acumulación térmica y no por el calendario, un modelo fenológico es capaz de calcular, a partir de la temperatura real, en qué estadio se encuentra la plaga en cada momento, incluso cuando permanece oculta al ojo humano.
Esta tecnología convierte la biología del insecto en información puramente operativa para el asesor técnico, permitiéndole saber con días de antelación:
- Cuándo aparecerán los primeros estadios L1–L2 antes de que el daño foliar sea visible.
- En qué fecha exacta se concentrará el pico máximo de eclosión.
- El momento preciso en que la población remanente entrará en fases biológicas resistentes.
Cambio de Paradigma Fitosanitario
Comparativa del impacto técnico y económico según el criterio de intervención en el lote.
- Disparador: Detección visual de daños por defoliación en el cultivo.
- Estado de la Plaga: Población dominada por estadios avanzados (L3 a L6).
- Condición Biológica: Larvas con máxima expresión enzimática detoxificadora y hábitos de refugio nocturno.
- Estrategia Química: Aplicaciones de «rescate» con dosis máximas y mezclas agresivas.
- Disparador: Alertas matemáticas basadas en la acumulación de Grados Día (GDD).
- Estado de la Plaga: Ventana biológica crítica de eclosión (Estadios L1 y L2).
- Condición Biológica: Cutícula fina permeable, detoxificación basal e insectos expuestos en el dosel.
- Estrategia Química: Uso eficiente de IGRs, diamidas o espinosinas sincronizadas con el MoA.
Beneficios Estratégicos del Enfoque Predictivo con Grados Día
1. Optimización del Retorno de Inversión (ROI)
Al aplicar exclusivamente en estadios L1–L2, cada euro invertido en fitosanitarios alcanza su máximo techo de rendimiento biológico, eliminando los costosos e inútiles «tratamientos de rescate» sobre larvas grandes.
2. Sostenibilidad del Negocio Agrícola
La predictividad permite reducir el número total de intervenciones químicas por ciclo, facilitando el cumplimiento de normativas de residuos e impulsando una producción alineada con las exigencias de la cadena agroalimentaria.
3. Planificación Logística Integral
Anticipar con días de antelación el pico de eclosión larvaria permite coordinar compras de stock con proveedores, gestionar agendas de operarios y calibrar la maquinaria con ventanas de tiempo óptimas, evitando compras reactivas de urgencia.
4. Preservación de Tecnologías Químicas
Aplicar de forma sincronizada con las directrices de IRAC frena la presión de selección de superresistencias metabólicas, salvaguardando la vida útil y efectividad de las moléculas clave (como diamidas y espinosinas) en la región.
Al anticipar la línea temporal del insecto, el manejo pasa de ser reactivo e ineficiente a preventivo, preciso y económicamente sostenible. Esta capacidad de anticipación matemática es precisamente donde FuturCrop aporta su mayor valor estratégico. Alinear las aplicaciones con la realidad biológica de la plaga no solo disminuye la presión de selección de resistencias y protege las herramientas químicas del mercado, sino que reduce el número total de tratamientos y maximiza la eficiencia de cada euro invertido en sanidad vegetal.
En última instancia, el desafío de Chrysodeixis includens recuerda un principio fundamental: la agronomía no es solo química, tecnología o experiencia acumulada; es, sobre todo, biología aplicada a decisiones concretas en el campo. Cuando la ingeniería de la aplicación se subordina a la predicción biológica, incluso la plaga más costosa se transforma en un problema bajo control.
Controla la Falsa Medidora como un Experto.
Sustituye la incertidumbre por datos fenológicos precisos. Con la Ficha de Suscripción de *Ch. includens*, obtienes acceso completo a alertas y seguimientos.
PROBAR GRATIS 7 DÍAS(Si decides continuar con la suscripción completa, cuesta solo 7 USD mensuales). Cancela cuando quieras.
*Los porcentajes de ahorro se basan en optimizaciones promedio de tratamientos fitosanitarios y reducción de pérdidas por daños masivos.*